|
|
Quiz Ch. 16 Quiz Ch. 17 |
Viruses: general principles. Bacteriophages |
|
|
|
Lecture Index | ||
|
|
Course Resources page |
Last revised: Wednesday, April 5, 2000
Ch. 16, 17 in Prescott et al, Microbiology, 4th Ed.Note: These notes are provided as a guide to topics the instructor hopes to cover during lecture. Actual coverage will always differ somewhat from what is printed here. These notes are not a substitute for the actual lecture!Copyright 2000. Thomas M. Terry
Every virus has two stages
- a dormant, particulate, transmissible stage called the virion stage
- an active, intracellular stage called the infectious stage
Virion Stage
- Virions are the transmissible state of a virus. Metabolically inert.
- Virion = "a piece of bad news wrapped up in a protein coat" (and/or a membrane)
- The "bad news" can be either DNA (double-stranded (ds) or single-stranded (ss)) or RNA (ds or ss); never both.
- The coat (also called viral shell or capsid) can be icosahedron (20-sided regular geometric shape common in many bacterial, animal, and plant viruses), sphere, cylinder, bullet-shaped, or amorphous shaped particle.
- Virions must be able to adhere and allow entry into some host cell(s). Also to survive outside of host cell environment.
- Some virions more hardy than others (e.g., hepatitis virus can withstand short periods of boiling; most virions are destroyed by this).
Infectious Stage
- When virus infects a cell, nucleic acid must be uncoated and gain access to metabolic machinery of cell.
- Virus life cycle is characterized by:
- attachment
- penetration, with entry of nucleic acid into cell
- early expression of virus genes (either directly by translation, if virus contains "+" RNA, or indirectly after transription and then translation)
- replication of virus nucleic acid
- synthesis of new virion components
- packaging and assembly of new virions
- exit from cell
- But note: some viruses can have variations on this theme: e.g., lysogeny (see below), retroviruses (see below), etc.
Measurement of viral growth
- Must grow virus on host cells to see anything. Can't grow virus without cells.
- To quantify viruses, need some way to get flat surface of growing cells, allow virus-infected cells to spread radially where present = plaque.

- In bacterial cells this is easy. Spread "lawn" of bacteria on plate, add diluted phage suspension or culture infected with phages. After 6-8 hours can see plaques in E. coli.
- In plant cells, can be easy. Example: Tobacco Mosaic Virus (TMV), make virus dilution, rub over surface of tobacco leaf. After leaf growth, can observe plaque areas.
- In animal cells, not so easy. In 1960's, standard assay was to inoculate chicken egg membranes of developing chick embryos, incubate for a week, cut open shell and count plaques on membrane in the air sac. Lots of work to get statistically reliable data!
- In 1970's tissue culture became a viable alternative. Animal cells are cultured as microbes in glass or plastic, use special medium that contains most of nutrients present in blood. Cells will spread as monolayer on surface, can count plaques after staining.
- Note: When counting plaques, report results as Plaque Forming Units, or PFU/ml,not as viruses/ml. Why? When counting viruses per unit volume under EM, often see far more than will cause plaques. (For some viruses only 1 in 100 or even 1000 particles will actually be detectable by plaque assay).
- Viral growth curve (in lab last week). Look at PFU/ml over time. See latent phase, then increase in PFU during viral release.
- View a short history of virology
Taxonomy of viruses
- Based mainly on Virion and Kingdom of host
- Use Host cell type (Animal viruses, plant viruses, etc.)
- Use Nucleic Acid type (ds DNA, ss DNA, ds RNA, ss RNA)
- Use + or - polarity of RNA. "+" is able to serve as mRNA. "-" is the complement of +, must function as template to make a complementary strand of + RNA before any translation can occur.
- Use virus coat morphology. Enveloped vs. non-enveloped viruses.
- Explore virus taxonomy
Virion Structure
See Lecture notes on "Virus structure"
- "Naked" viruses
- Helical viruses
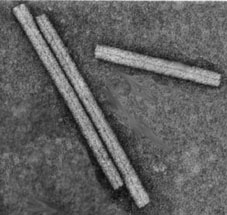
- Tobacco mosaic virus (TMV) is an example of a virus with helical symmetry.
- A helical array of identical protein subunits surrounds an RNA molecule
TMV electron micrograph: image from University of Leicester
- Icosahedral viruses
- built from icosahedral (20-sided) assemblies of protein subunits.
- View animation of icosahedron shape (from Tulane University)
- Icosahedral shape is the minimum free energy structure for producing a shell of equivalently bonded identical structures.
- The simplest icosahedral capsids are built up by using 3 identical subunits to form each triangular face, thereby requiring 60 identical subunits to form a complete capsid. A few simple virus particles are constructed in this way, e.g. bacteriophage �X174.
- Most icosahedral viruses have more than 60 subunits, usually some multiple N times 60. N (called the triangulation number) can have values of 1, 3, 4, 7, 9, 12, and more.
- Visual example: adenovirus diagram
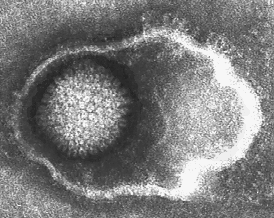
- "Enveloped" viruses
- "Naked" viruses require host death so viruses can be released. This may be wasteful, and may cause premature death of host cell.
- Alternative strategy: shed virus particles by budding out, continued release from cell membrane. Cell does not die (immediately), continues to serve as factory for virus assembly and release. Virus typically acquires a coating of host cell membrane, modified to include virus-specific proteins. This is the "envelope". Virus may have additional protein coats (often icosahedral) inside the envelope.
Enveloped Herpes virus, by Linda Stannard
- Eventually host cell is too depleted to survive. Can see evidence of this as "cytopathic effect" (CPE). Cell then dies.
- Examples of enveloped viruses include:
- Retrovirus, including HIV
- Paramyxovirus, including influenza
- Rhabdovirus, including rabies
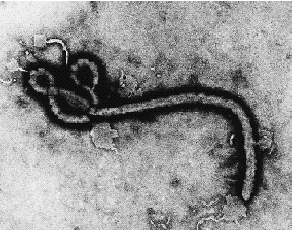
- Filovirus. Although very "hot" in the news, these viruses are very poorly characterized because of their extreme pathogenicity. They are class IV pathogens, meaning they can only be cultured in total containment facilities, of which there are only two in the U. S. They are thought to be enveloped viruses with - RNA genomes.
Ebola virus image by Dr. Frederick A. Murphy, University of California, DavisVirus Genomes
- Rule of Thumb: to estimate # of virus proteins, look at size of viral DNA or RNA. For each 1000 base pairs, can guess the existence of 1 protein
- "typical" gene has 300-400 amino acids = ~ 1000 base pairs = 1 kbp (= 1 protein)
- small virus: SV40 => 5000 base pairs = 5 kbp ~ 5 proteins
- large virus: T4 => 200 kbp ~ 100-200 proteins
- by comparison, E. coli: 4000 kbp
- For more detailed information, visit the structure and complexity of viral genomes from the University of Leicester
Bacterial Viruses = Phages
- Animate Stages of Lytic virus infection
- For further information beyond the three phages studied here, visit Bacteriophages, from University of Leicester, U. K.
Case Study #1: small RNA phage MS2 ---> lytic infection
- MS2 is one of the simplest viruses known; isolated many times, different names (R17, F2, MS2, Q-beta, etc.). See class handout, also figure 8.15 and related text.
- Virion is only 250 Angstroms in diameter, has 3500 nucleotides of ss RNA (3 genes). Virion contains two proteins: coat protein (many copies), and "A" protein (one copy/virion). A is needed for adsorption and entry.
- Virus can infect E. coli cell, produce 5000-10,000 progeny in 30-60 min at 37 deg C. How can it do so much with so few genes?
- Life cycle:
- virion adsorbs to host cell, RNA gets in (exactly how not clear)
- RNA strand attaches to host ribosomes, has "AUG" start codon, gets transcribed (this is a "+" RNA)
- First protein decoded is Replicase: binds to +RNA, makes -RNA copies.
- Once -RNA copies appear, replicase preferentially binds to them (10 to 1 preference for - vs + as template to copy), makes mostly + RNA strands.
- Coat protein is synthesized, accumulates in quantity, self-assembles around RNA to form new virions.
- Lytic step releases progeny phage.
Case study #2: large phage T4 ---> lytic infection
- View EM of phage T4
- Virion has ds DNA (170 kbp), over 135 genes described. Icosahedral head, tail with contractile sheath, lysozyme at tip of tail, fibers, base plate, etc. Very complex morphology. (see class handout; also pp. 270-272 in text)
- View T4 morphology
- Virus DNA is injected, easily enters host. DNA has promoter sites that look just like host ("TATA" boxes, etc.)
- View diagram of T4 DNA entry
- RNA polymerase of host sees DNA, transcribes mRNA. This gets translated---> "early proteins".
- Genes grouped into early (uses host sigma) and late (uses new T4 sigma). One of the early proteins specifically blocks host sigma.
- Examples of "early gene" function
- endonuclease attacks host DNA
- new sigma factor to recognize only phage specific promoters (Not TATA; something altogether different)
- enzymes to produce new kind of nucleotide found in virus DNA: hydroxymethyl cytosine. (T4 DNA does not contain normal cytosine)
- enzymes for DNA replication. T4 phages use different origin than on host chromosome; don't produce theta structures; instead, produce long strings of repeated phage DNA = "concatomers". DNA replication occurs on linear molecule. Results in concatamer of repeated genes. At packaging time, enzyme cuts DNA into head sized units (includes some repeat at each end of DNA)
- new t-RNAs
- new ribosomal proteins (work better on T4 messages)
- After early genes have been expressed, cell switches to "late genes"
- Examples of "late gene" function:
- phage head and tail assembly
- packaging of phage DNA into phage heads
- production of T4 lysozyme, and lysis of cell
Case Study #3: lysogenic phage Lambda ---> lytic or lysogenic infection
- View animation of stages in Phage lambda infection and induction
- Lambda has ds DNA, injected into cell as linear molecule. Enough DNA for ~ 55 proteins. (see section 8.12 in text, also class handout)
- View image of phage lambda
- Phage has two possible outcomes: lytic infection or lysogeny = "silent" partnership with cell, virus DNA becomes integrated into host chromosome and gets duplicated and passed on to all cell offspring.
- DNA has "sticky ends". ~20 bases at each end are single stranded. The two ends are complementary, DNA can circularize once in cell. DNA ligase can seal two ends to make covalently closed circle.
- Lambda has 3 promoter sites. Two of these allow transcription of lytic genes. Other promoter leads to transcription of a repressor protein (Lambda repressor) that can bind to the two lytic promoters, block all lytic genes. Repressor does not block its own promoter, so cell continues to synthesize small quantity of repressor (~10-20 copies/cell).
- One early lambda protein is Integrase; causes specific recombination event at region where both Lambda DNA and host DNA have same 13 base pairs. (homologous sequence).
- If Lambda repressor is expressed before transcription of late lytic pathway genes occurs, then Lambda remains in host DNA indefinitely, gets replicated just like host genes.
- Induction: under "nasty" environmental conditions where DNA damage occurs (e.g. UV light or certain chemicals), can stop repressor synthesis. If this occurs, lytic promoters are no longer blocked, lytic genes get transcribed and translated, and cell becomes phage factory, leads to lytic production of lambda viruses.
Bacterial defenses against infection
Cell surfaces: possibilities of mutation
- Virus must attach to some specific cell surface protein or polysaccharide. But these are specified by genes, and genes can mutate. In population, will always find some variant strains with slightly different cell surfaces, may not bind virus well.
- When phage first discovered, thought this could be effective weapon against bacterial disease. But frequency of resistant bacterial strains was too high, any given strain of virus quickly became useless as resistant survivors propagated.
Nucleases: endo- and exo-DNases and RNases
- All bacteria seem to have nucleases that can attack DNA (called DNases) and RNA (called RNases).
- Exoenzymes attack free 5' or 3' ends of DNA, RNA molecules. Bacteria are protected since DNA (and plasmids) are always circular. RNases are present, and in fact destroy mRNA eventually (bacteria are always making new RNAs, very responsive to enviroment changes).
- Endonucleases are potentially lethal weapons. Called restriction enzymes. Attack at specific sequence: e.g., in E. coli, enzyme called EcoRI will attack any sequence with 5' G-A-A-T-T-C 3' (cuts DNA between G and A).
- Why doesn't this kill cell? Because cell also has a second enzyme, called modification enzyme, that protects all host DNA sequences of this type. Typically adds a methyl (-CH3) group to one base at the cutting site. The methylated base is modified, and protected from the restriction enzyme. When foreign DNA comes into cell (e.g. virus DNA), if restriction site if present it will be cut and ----- requiem for the virus!
The importance of Restriction Enzymes
Restriction enzymes are responsible for the genetic revolution. They make reproducible, specific cuts with surgical precision. Major industry has emerged in biochemical supply companies to harvest bacteria, purify restriction enzymes, and sell these to research and applied industries. Big $$$$$$$.
Take a Self-Quiz on Ch. 16 material
Take a Self-Quiz on Ch. 17 material
Return to Lecture Index
Return to MCB 229 Course Resources page
{kind=link}
{kind=link}
{kind=link}