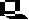
Golgi Apparatus and Dictyosomes | Lysosomes |Mitochondria | Plastids | Cell Movement
Shape of cells is varied with some, such as neurons, being longer than they are wide and others, such as parenchyma (a common type of plant cell) and erythrocytes (red blood cells) being equidimensional.
Size of cells is also related to their functions. Eggs (or to use the latin word, ova) are very large, often being the largest cells an organism produces (although not in all cases, some organisms put all their eggs in a large basket while others produce a plethora of small eggs). The large size of many eggs is related to the process of development that occurs after the egg is fertilized, when the contents of the egg (now termed a zygote) are used in a rapid series of cellular divisions, each requiring tremendous amounts of energy that is available in the zygote cells. Later in life the energy must be acquired, but at first a sort of inheritance/trust fund of energy is used.
The cell membrane functions as a semi-permeable barrier, allowing a very few molecules across it while fencing the majority of organically produced chemicals inside the cell. Electron microscopic examinations of cell membranes have led to the development of the lipid bilayer model (also referred to as the fluid-mosaic model). The most common molecule in the model is the phospholipid, which has a polar (hydrophilic) head and two nonpolar (hydrophobic) tails. These phospholipids are aligned tail to tail so the nonpolar areas form a hydrophobic region between the hydrophilic heads on the inner and outer surfaces of the membrane. This layering is termed a bilayer since an electron microscopic technique known as freeze-fracturing is able to split the bilayer.
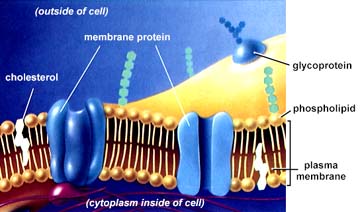
Diagram representing the cell membrane. The above image is from http://www.biosci.uga.edu/almanac/bio_103/notes/may_15.html.
Cell Membranes from Opposing Neurons (TEM x436,740). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Cholesterol is another important component of cell membranes embedded in the hydrophobic areas of the inner (tail-tail) region. Most bacterial cell membranes do not contain cholesterol.
Proteins are suspended in the inner layer, although the more hydrophilic areas of these proteins "stick out" into the cells interior and outside of the cell. These proteins function as gateways that will, in exchange for a price, allow certain molecules to cross into and out of the cell. These integral proteins are sometimes known as gateway proteins. The outer surface of the membrane will tend to be rich in glycolipids, which have their hydrophobic tails embedded in the hydrophobic region of the membrane and their heads exposed outside the cell. These, along with carbohydrates attached to the integral proteins, are thought to function in the recognition of self.
The contents (both chemical and organelles)of the cell are termed protoplasm, and are further subdivided into cytoplasm (all of the protoplasm except the contents of the nucleus) and nucleoplasm (all of the material, plasma and DNA etc. within the nucleus).
Not all living things have cell walls, most notably animals and many of the more animal-like Protistans. Plant cells have a variety of chemicals incorporated in their cell walls. Cellulose is the most common chemical in the plant primary cell wall. Some plant cells also have lignin and other chemicals embedded in their secondary walls. The cell wall is located outside the cell membrane. Plasmodesmata are connections through which cells communicate chemically with each other through their thick walls. Fungi and many protists have cell walls although they do not contain cellulose, rather a variety of chemicals (chitin for fungi).
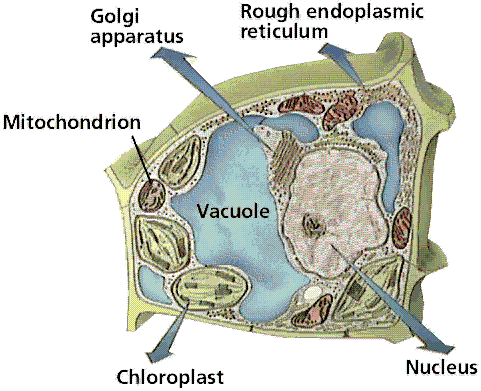
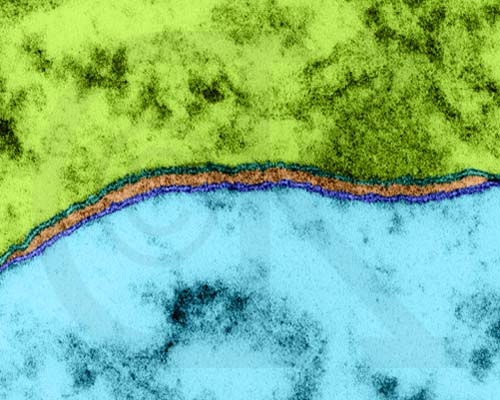
Structure of a typical plant cell. Image from W.H. Freeman and Sinauer Associates, used by permission.
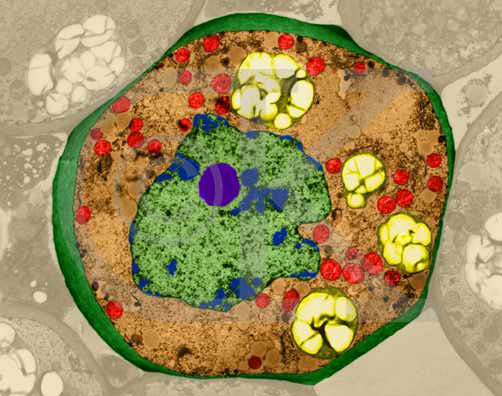
Lily Parenchyma Cell (cross-section) (TEM x7,210). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission. Note the large nucleus and nucleolus in the center of the cell, mitochondria and plastids in the cytoplasm.
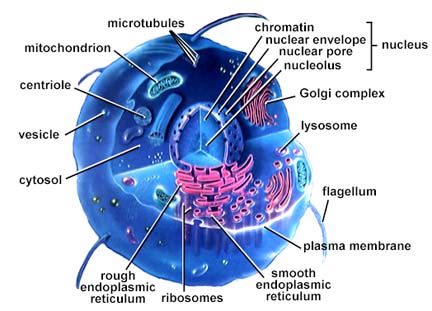
Structure of an animal cell. The above image is from http://www.biosci.uga.edu/almanac/bio_103/notes/may_15.html.
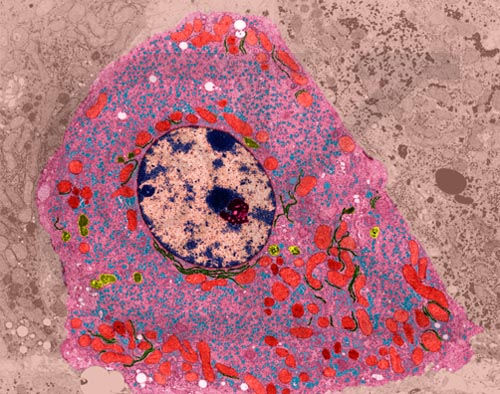
Liver Cell (TEM x9,400). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
The nucleus (follow link to the MIT Hypertextbook page) occurs only in eukaryotic cells, and is the location of the majority of different types of nucleic acids. Van Hammerling's experiment (click here for a diagram) showed the role of the nucleus in controlling the shape and features of the cell. Deoxyribonucleic acid, DNA, is the physical carrier of inheritance and with the exception of plastid DNA (cpDNA and mDNA, see below) all DNA is restricted to the nucleus. Ribonucleic acid, RNA, is formed in the nucleus by coding off of the DNA bases. RNA moves out into the cytoplasm. The nucleolus is an area of the nucleus (usually 2 nucleoli per nucleus) where ribosomes are constructed.
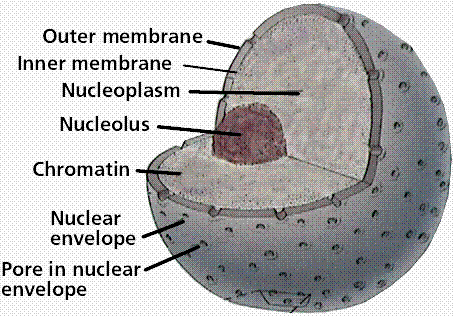
Structure of the nucleus. Note the chromatin, uncoiled DNA that occupies the space within the nuclear envelope. Image from W.H. Freeman and Sinauer Associates, used by permission. Image from W.H. Freeman and Sinauer Associates, used by permission.
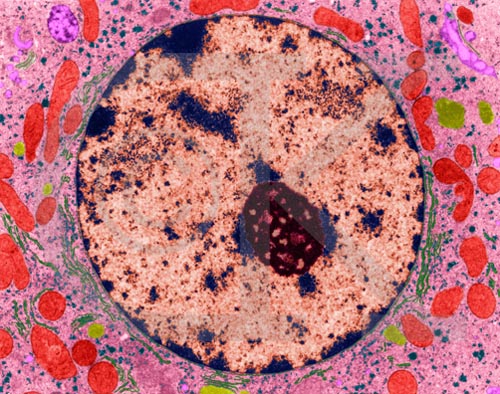
Liver cell nucleus and nucleolus (TEM x20,740). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission. Cytoplasm, mitochondria, endoplasmic reticulum, and ribosomes also shown.
The nuclear envelope is a double-membrane structure. Numerous pores occur in the envelope, allowing RNA and other chemicals to pass, but the DNA not to pass.
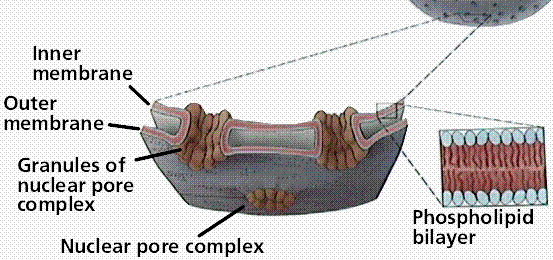
Structure of the nuclear envelope and nuclear pores. Image from W.H. Freeman and Sinauer Associates, used by permission. Image from W.H. Freeman and Sinauer Associates, used by permission.
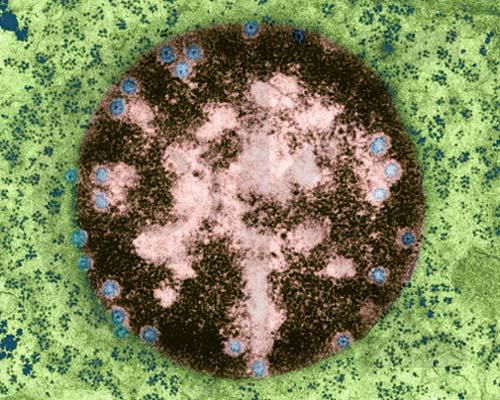
Nucleus with Nuclear Pores (TEM x73,200). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission. Cytoplasm also contains numerous ribosomes.
The cytoplasm was defined earlier as the material between the plasma membrane (cell membrane) and the nuclear envelope. Fibrous proteins that occur in the cytoplasm, referred to as the cytoskeleton maintain the shape of the cell as well as anchoring organelles, moving the cell and controlling internal movement of structures. Microtubules function in cell division and serve as a "temporary scaffolding" for other organelles. Actin filaments are thin threads that function in cell division and cell motility. Intermediate filaments are between the size of the microtubules and the actin filaments.
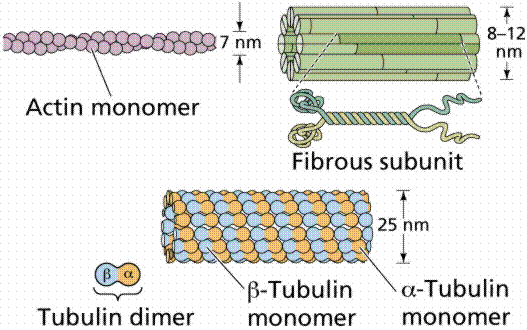
Actin and tubulin components of the cytoskeleton. Image from W.H. Freeman and Sinauer Associates, used by permission.
Vacuoles are single-membrane organelles that are essentially part of the outside that is located within the cell. The single membrane is known in plant cells as a tonoplast. Many organisms will use vacuoles as storage areas. Vesicles; are much smaller than vacuoles and function in transport within and to the outside of the cell.
Ribosomes are the sites of protein synthesis. They are not membrane-bound and thus occur in both prokaryotes and eukaryotes. Eukaryotic ribosomes are slightly larger than prokaryotic ones. Structurally the ribosome consists of a small and larger subunit. Biochemically the ribosome consists of ribosomal RNA (rRNA) and some 50 structural proteins. Often ribosomes cluster on the endoplasmic reticulum, in which case they resemble a series of factories adjoining a railroad line. Click here for Ribosomes (More than you ever wanted to know about ribosomes!)
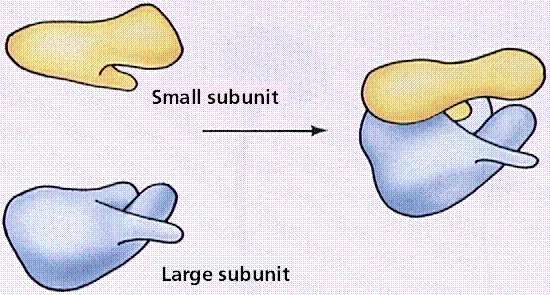
Structure of the ribosome. Image from W.H. Freeman and Sinauer Associates, used by permission.
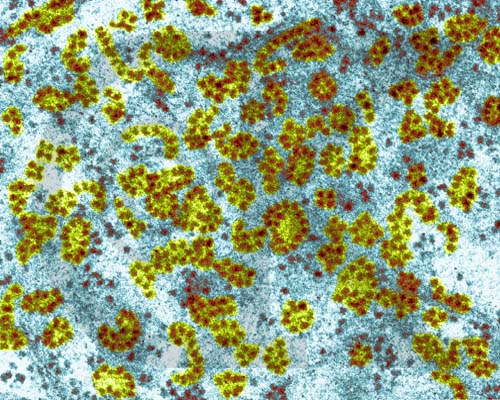
Ribosomes and Polyribosomes - liver cell (TEM x173,400). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Endoplasmic reticulum is a mesh of interconnected membranes that serve a function involving protein synthesis and transport. Rough endoplasmic reticulum (Rough ER) is so-named because of its rough appearance due to the numerous ribosomes that occur along the ER. Rough ER connects to the nuclear envelope through which the messenger RNA (mRNA) that is the blueprint for proteins travels to the ribosomes. Smooth ER; lacks the ribosomes characteristic of Rough ER and is thought to be involved in transport and a variety of other functions. Click here to view the WWW Cell Biology Course page on the endoplasmic reticulum.
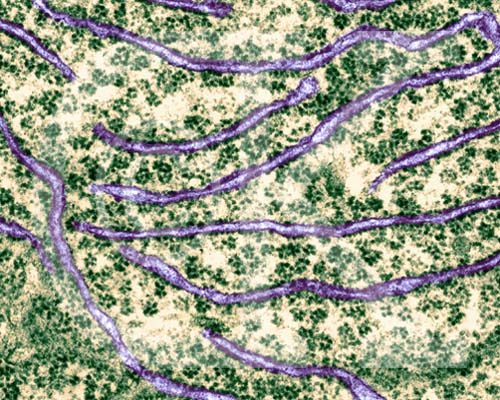
The endoplasmic reticulum. Rough endoplasmic reticulum is on the left, smooth endoplasmic reticulum is on the right. Image from W.H. Freeman and Sinauer Associates, used by permission.
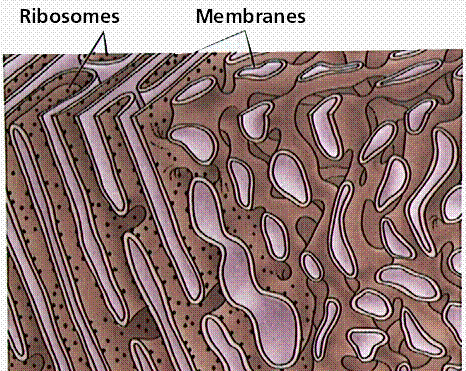
Rough Endoplasmic Reticulum with Ribosomes (TEM x61,560). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Golgi Complexes (follow the link to the MIT Hypertextbook page on Golgi) are flattened stacks of membrane-bound sacs. They function as a packaging plant, modifying vesicles from the Rough ER. New membrane material is assembled in various cisternae of the golgi.
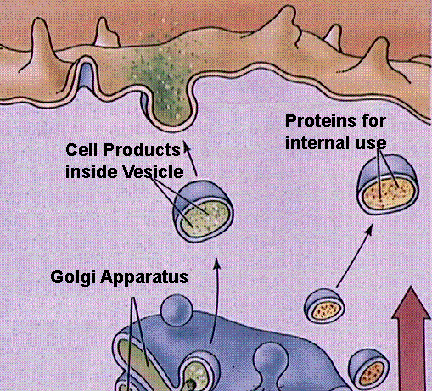
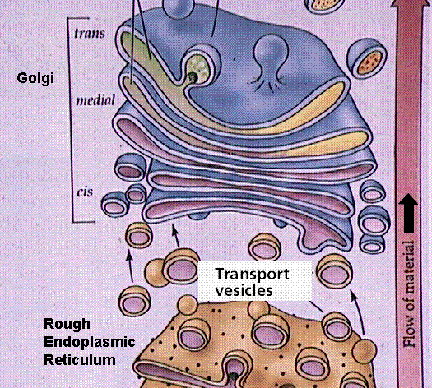
Structure of the Golgi apparatus and its functioning in vesicle-mediated transport. Images from W.H. Freeman and Sinauer Associates, used by permission.
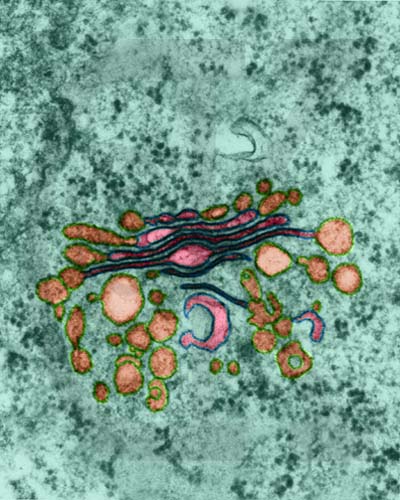
Golgi Apparatus in a plant parenchyma cell from Sauromatum guttatum (TEM x145,700). Note the numerous vesicles near the Golgi. This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
Lysosomes are relatively large vesicles formed by the Golgi. They contain hydrolytic enzymes that could destroy the cell. Lysosome contents function in the extracellular breakdown of materials.
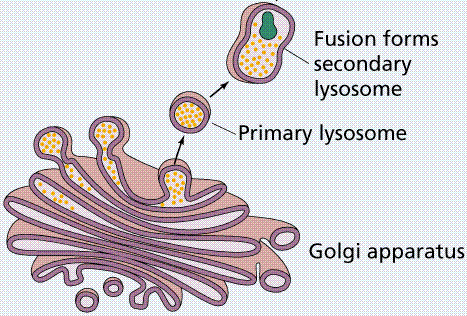
Role of the Golgi in forming lysosomes. Image from W.H. Freeman and Sinauer Associates, used by permission.
Mitochondria (follow link to the mitochondria drawing from the MIT Hypertext) contain their own DNA (termed mDNA) and are thought to represent bacteria-like organisms incorporated into eukaryotic cells over 700 Ma ago (perhaps as far back as 1.5 billion years ago). They function as the sites of energy release (following glycolysis in the cytoplasm) and ATP formation (by chemiosmosis). The mitochondrion has been termed the powerhouse of the cell. Mitochondria are bounded by two membranes. The inner membrane folds into a series of cristae, which are the surfaces on which ATP is generated.Click here to view the WWW Cell Biology page about mitochondria.
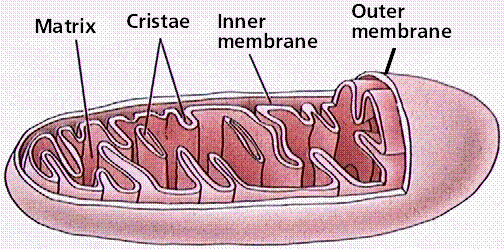
Structure of a mitochondrion. Image from W.H. Freeman and Sinauer Associates, used by permission.

Muscle Cell Mitochondria (TEM x190,920). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission.
During the 1980s, Lynn Margulis proposed the theory of endosymbiosis to explain the origin of mitochondria and chloroplasts from permanent resident prokaryotes. According to this idea, a larger prokaryote (or perhaps early eukaryote) engulfed or surrounded a smaller prokaryote some 1.5 billion to 700 million years ago.
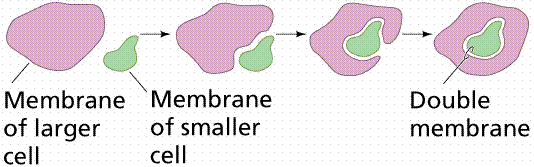
The basic events in endosymbiosis. Image from W.H. Freeman and Sinauer Associates, used by permission.
Instead of digesting the smaller organisms the large one and the smaller one entered into a type of symbiosis known as mutualism, wherein both organisms benefit and neither is harmed. The larger organism gained excess ATP provided by the "protomitochondrion" and excess sugar provided by the "protochloroplast", while providing a stable environment and the raw materials the endosymbionts required. This is so strong that now eukaryotic cells cannot survive without mitochondria (likewise photosynthetic eukaryotes cannot survive without chloroplasts), and the endosymbionts can not survive outside their hosts. Nearly all eukaryotes have mitochondria. Mitochondrial division is remarkably similar to the prokaryotic methods that will be studied later in this course. A summary of the theory is available by clicking here.
Plastids are also membrane-bound organelles that only occur in plants and photosynthetic eukaryotes.
Chloroplasts are the sites of photosynthesis in eukaryotes. They contain chlorophyll, the green pigment necessary for photosynthesis to occur, and associated accessory pigments (carotenes and xanthophylls) in photosystems embedded in membranous sacs, thylakoids (collectively a stack of thylakoids are a granum [plural = grana]) floating in a fluid termed the stroma. Chloroplasts contain many different types of accessory pigments, depending on the taxonomic group of the organism being observed.
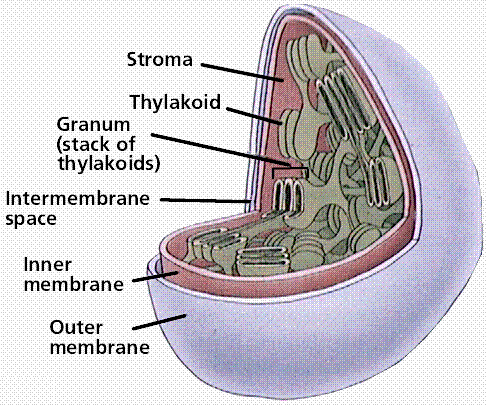
Structure of the chloroplast. Image from W.H. Freeman and Sinauer Associates, used by permission.
Like mitochondria, chloroplasts have their own DNA, termed cpDNA. Chloroplasts of Green Algae (Protista) and Plants (descendants of some Green Algae) are thought to have originated by endosymbiosis of a prokaryotic alga similar to living Prochloron (Prochlorobacteria). Chloroplasts of Red Algae (Protista) are very similar biochemically to cyanobacteria (also known as blue-green bacteria [algae to chronologically enhanced folks like myself :)]). Endosymbiosis is also invoked for this similarity, perhaps indicating more than one endosymbiotic event occurred.
Leukoplasts store starch, sometimes protein or oils.
Chromoplasts store pigments associated with the bright colors of flowers and/or fruits.
Cell movement; is both internal, referred to as cytoplasmic streaming and external, referred to as motility. Internal movements of organelles are governed by actin filaments. These filaments make an area in which organelles such as chloroplasts can move. Internal movement is known as cytoplasmic streaming. External movement of cells is determined by special organelles for locomotion.
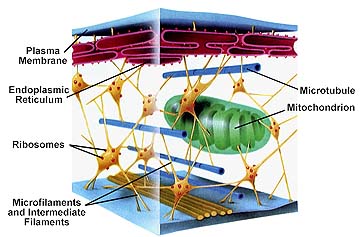
The cytoskeleton.
Cilia and flagella are similar except for length, cilia being much shorter. They both have the characteristic 9 + 2 arrangement of microtubules.
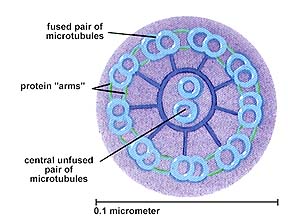
The 9+2 arrangement of microtubules in a flagellum or cilium.

Cilia from an epithelial cell in cross section (TEM x199,500). This image is copyright Dennis Kunkel http://www.pbrc.hawaii.edu/~kunkel/gallery, used with permission. Again, note the 9 + 2 arrangement of cilia.
Flagella work as whips pulling (as in Chlamydomonas or Halosphaera) or pushing (dinoflagellates, a group of single-celled Protista) the organism through the water. Cilia work like oars on a viking longship (Paramecium has 17,000 such oars covering its outer surface).

Movement of cilia and flagella. Image from W.H. Freeman and Sinauer Associates, used by permission.
Pseudopodia are used by many cells, such as Amoeba, Chaos (Pelomyxa) and human leukocytes (white blood cells). These are not structures as such but rather are associated with actin near the moving edge.

Formation and functioning of a pseudopod by an amoeboid cell. Image from W.H. Freeman and Sinauer Associates, used by permission.
These learning objectives are taken from my Biology for Nonmajors class (BIO 102). I have tried to add a link to each that will direct you to a part of this chapter or another website that will facilitate your completion of the objective.
Back to Table of Contents | Go to TRANSPORT IN AND OUT OF CELLS
Email: mj.farabee@emcmail.maricopa.edu![]()
Last modified: 2000/01/09:09:46:48
The URL of this page is: gened.emc.maricopa.edu/bio/BIO181/BIOBK/BioBookCELL2.html
{kind=link}
{kind=link}
{kind=link}