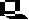
Organization of the Animal Body | Evolution and Classification of Animals
Trends in Animal Evolution | Sponges: The Phylum Porifera
Tissues: Jellyfish, Corals and Sea Anemones
Bilateral Symmetry and Cephalization: Phylum Platyhelminthes
The Phylum Nemertea: Ribbon Worms | The Phylum Rotifera
The Tube-within-a-tube Body Plan: Phylum Nematoda | Links
Animals are characteristically multicellular heterotrophs whose cells lack cell walls. At some point during their lives, animals are capable of movement. In the most commonly encountered animals, this stage is the adult, although some animals (corals) have sessile (nonmobile) adult phases and mobile juvenile forms. Animal and plant evolutionary history both show the development of multicellularity and the move from water to land (as well as secondary adaptation back to water).
Animals developed external or internal skeletons to provide support, skin to prevent or lessen water loss, muscles that allowed them to move in search of food, brains and nervous systems for integration of stimuli, and internal digestive systems.
Most animals have a life cycle with a pre-adult stage, a predominance of the diploid stage, and a series of embryonic developmental stages.
The above image is from http://www.biosci.uga.edu/almanac/bio_103/notes/apr_4.html.
Animals probably evolved from marine protists, although no group of protists has been identified from an at-best sketchy fossil record for early animals. Cells in primitive animals show similarities to collared choanoflagellates as well as pseudopodia-producing amoeboid cells.
Multicellular animal fossils and burrows (presumably made by multicellular animals) first appear 700 million years ago, during the late precambrian time (the Proterozoic). All known Proterozoic animal fossils had soft body parts: no shells or hard (and hence preservable as fossils) parts. Learn more about these early animal fossils at Learning About the Vendian Animals.
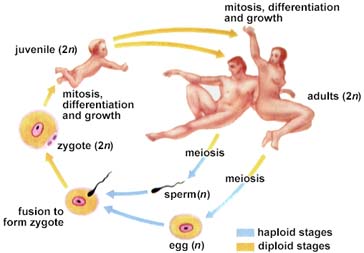
First appearances and relative diversity (width of shaded area) for major groups of animals. Image from W.H. Freeman and Sinauer Associates, used by permission.
Beginning 570 million years ago, the early Cambrian Period, animals with external skeletons appeared in abundance. This suddenn appearance of fossils is in fact used to define the beginning of the Cambrian (named after Cambria, an ancient name for Wales). These external skeletons were hard enough to be more readily preserved than the soft-bodied forms that had dominated early animal evolution.
Modern animals are classified into between 30 and 35 phyla: all major modern phyla were present at the beginning of the Cambrian, along with a great variety of now-extinct phyla recorded in the Burgess Shale (Cambrian) in Canada. Of the animal phyla, scientists consider nine major invertebrate phyla and the chordates to be of major importance in terms of biological diversity. While all major animal phyla are represented by Cambrian fossils, reconstructing fossil history is extremely difficult since earlier, soft-bodied animals did not fossilize well. Consequently, evolutionary relationships have been established for the most part on a studies of living (extant) animal anatomy.
Most animals have a body plan best described as a "tube-within-a-tube". This plan calls for two openings: one for food to enter the body (mouth), one for wastes to leave the body (anus). Animals with the "tube-within-a-tube" plan are 10% more efficient at digesting and absorbing their food than animals with the sac-like body plan. The tube-within-a-tube plan allows specialization of parts along the tube. The sac-like body plan has only one opening for both food intake and waste removal. Sac-like body plan animals do not have tissue specialization or development of organs.
Many, but not all, animals have three tissue layers as they develop embryologically: the endoderm, mesoderm, and ectoderm. Some animals, most notably sponges, lack these tissue layers. Cnidarians (coral and jellyfish) have only two of these layers. Flatworms, ribbon worms, etc. all have three tissue layers. Humans are triploblastic.
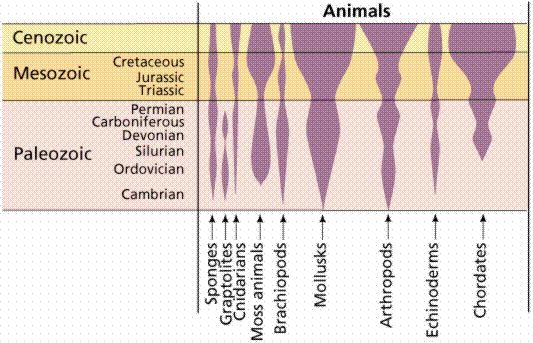
Asymmetrical animals (sponges) have no general body plan or axis of symmetry that divides the body into mirror-image halves. Radially symmetrical animals (such as coral and jelly fish) have body parts organized about a central axis and tend to be cylindrical in shape. Radially symmetrical animals often are (for some part of their life) nonmotile (sessile), and live attached to a substrate. Radial symmetry allows animals to reach out in all directions from one center. Bilaterally symmetrical animals (such as humans and fish) have only a single plane of symmetry that produces mirror halves. Bilaterally symmetrical animals tend to be active and to move forward at an anterior end, which eventually led to concentration of sensory organs in the anterior end, or head.
Different body plans. There are no planes of symmetry in the asymmetrical form, while the radial form has numerous planes. Bilateral organisms, like humans, have only one plane of symmetry, extending from the head to the toe. Bilateral symmetry is an important step in development of a head and concentartion of sensory organs in that head.
Acoelomate animals (like flatworms and flukes) do not have a coelom (or body cavity) produced during preadult development. Pseudocoelomate animals (like roundworms) have a body cavity but it does not develop from splitting of the mesoderm. Coelomate animals (humans, fish, shrimp) have a body cavity lined with mesoderm cells.
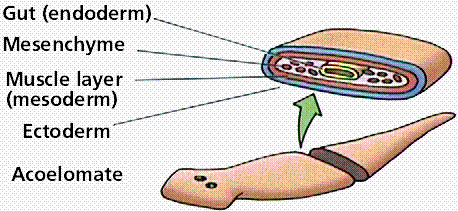
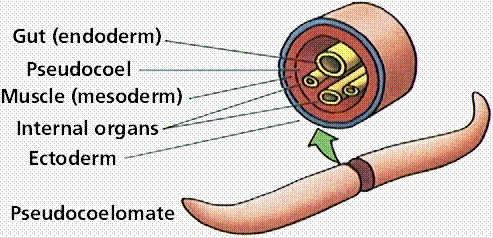
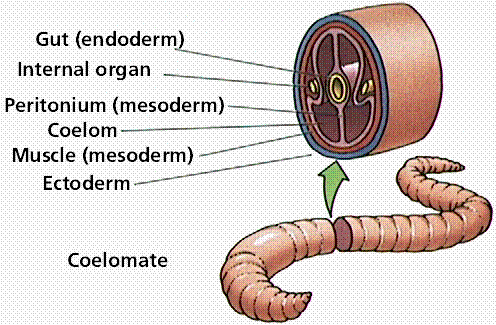
Three body cavity styles in modern animals.
Coelomates are either protostomes or deuterostomes. Protostomes (first mouth) are coelomates whose embryonic development shows a blastopore associated with a mouth. Deuterostomes (second mouth) are coelomates whose embryonic development shows a blastopore associated with an anus, with a second opening forming the mouth (hence "second mouth"). Vertebrates are deuterostomes.

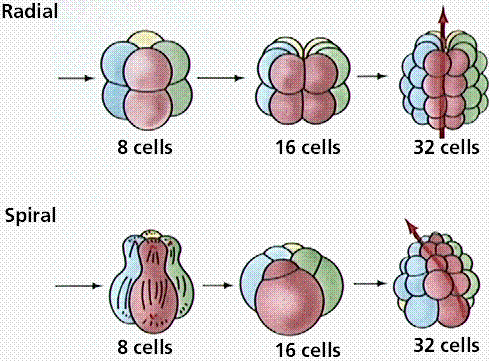
Differences in cleavage between the embryos of protostomes and deuterostomes.
Some animals have their bodies divided into segments. This allows them to specialize certain segments, such as for antennae, eyes, claws, etc. Humans, insects, and earthworms are examples of segmented animals.
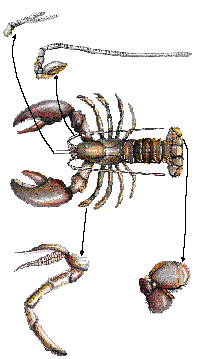
Examples of body segments as seen in a crayfish. Segmentation of the body allows development of various specialized limbs, such as antennae, pincers, walking legs, swimming legs, and feeding appendages.
The Phylum Porifera ("pore-bearing") contains approximate 5,000 species of sponges. These asymmetrical animals have sac-like bodies that lack tissues, and are usually interpreted as representing the cellular level of evolution. Cells from fragmented sponges can reorganize/regenerate the sponge organism, something not possible with animals that have tissues. Most zoologists consider sponges as offshoots that represent an evolutionary dead-end. Sponges are aquatic, largely marine animals, with a great diversity in size, shape, and color.
Modern sponges greatly resemble some fossil Cambrian sponges. Sponges may have evolved from a colonial protozoan. There are no true tissues in sponges: merely specialized cell layers. Epidermal cells in sponges line the outer surface. Collar cells line the inner cavity. Beating collar cells produce water currents that flow through pores in sponge wall into a central cavity and out through an osculum, the upper opening. A 10 cm tall sponge will filter as much as 100 liters of water a day. Amoeboid cells occupy the "inner" layer, along with hardened structures known as spicules.
Sponges feed by drawing water into the body through a network of pores (hence the name porifera, pore-bearer) and passing it out through the large opening (osculum) at one end of the body.
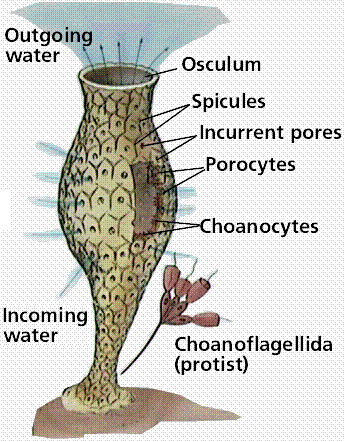

Organization of the sponge body. Note the lack of tissues. Sponges demonstrate the cellular level of organization. Also notice the resemblance between the collared choanoflagellate cells in the top drawing, and the collar cells in the sponge.
Sponges can reproduce asexually (by budding or from fragments) or sexually. Sponges produce eggs and sperm that are released into a central cavity of the sponge, in which the zygote develops into a ciliated larva. The larval stage is able to move about while the adult is stationary.
Image of some calcareous vase sponges belomnging to the genus Scypha. Image from http://128.128.172.102/SPECIMENS/phylum.taf?function=detail&ID=2&branch=image.
The fossil record of sponges has been at times quite good. The oldest sponges date from the precambrian. One early example of fossil sponges are the archaeocyathids, one of the first reef-building animals. Archaeocyathids evolved and went extinct before then end of the Cambrian Period. Cladistic analysis by J. Reitner in 1990 suggests archaeocyathids are properly placed in the Phylum Porifera instead of having their own phylum. Living sponges fall into three groups: the calcareous, glass, and demosponges.
The Phylum Cnidaria contains 10,000 species characterized by adult bodies having radial symmetry. Cnidarians are aquatic, mostly all marine. Cnidarians have only the ectoderm and endoderm tissue layers. Members of this phylum are characterized by stinging cells, that eject a barbed thread and possibly a toxin. Only cnidaria have cnidocytes, a specialized cell that contains a nematocyst, a fluid-filled capsule containing a long, spirally coiled hollow thread. When the trigger of the cnidocyte is touched, the nematocyst is discharged. Some threads merely trap a prey or predator, while others have spines that penetrate and inject paralyzing toxins.

Cnidocyte/nematocyst in a cnidarian. These stinging cells allow the animal to capture small prey, as well as offer some defense if attacked.
Two body forms occur: a mobile medusa and a sessile (not mobile) polyp. Both forms have tentacles arranged around an opening into the two-layered sac-like body. The inner tissue layer (derived from endoderm) secretes digestive juices into the gastrovascular cavity, which digests food and circulates nutrients. Muscle fibers occur at the base of the epidermal and gastrodermal cells. Nerve cells located below epidermis near the mesoglea interconnect and form a nerve net throughout the body. Cnidarians have both muscle fibers and nerve fibers, making these animals capable of directional movement. The nerve net allows transmission of messages in more than one direction, possibly an advantage in a radially symmetrical animal, while contraction of muscle fibers (under control of the nerve fibers) allows for movement.
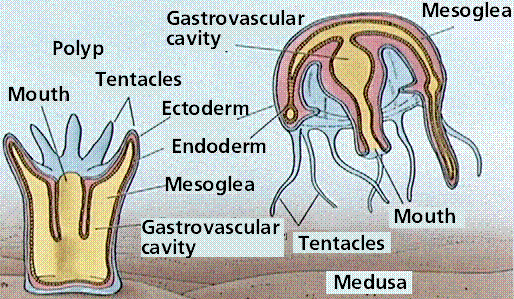
Body types in a typical cnidarian.
Cells are organized into tissues. The adult in most species is radially symmetrical. The typical cnidarian life cycle involves both sexual and asexual reproduction. A bilaterally symmetrical larva known as a planula, develops from a zygote. The planula settles in an appropriate location and grown into the adult polyp, that reproduces asexually to form medusae. Each medusa develops gonads and forms gametes.
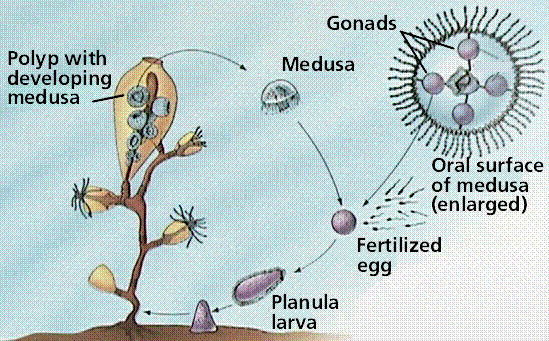
Life cycle of a typical cnidarian. In esswence we see an "alternation of generations" between the sessile polyp phase and the mobile medusa phase. However, unlike plants, both phases are diploid.
Cnidarians are divided into a number of taxonomic classes.
Sea anemones are solitary polyps 5-100 mm in height and 5-200 mm in diameter or larger. They are often brightly colored and look like flowers on the seafloor. Their thick, heavy body rests on a pedal disk and supports an upward-turned mouth surrounded by hollow tentacles. Sea anemones feed on various invertebrates and fish. They attach to a variety of substrates, or may be mutualistic with hermit crabs, living attached to crab's shell.
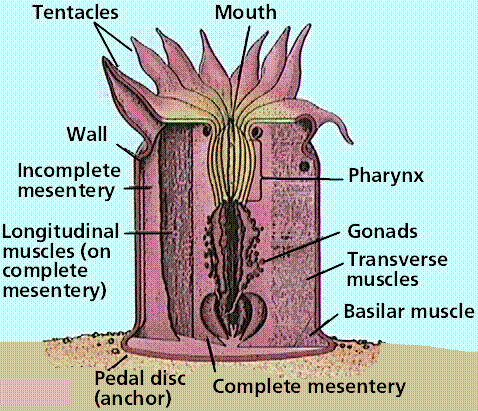
Anatomy of a polyp. These animals have an almost plant-like appearance, being anchored in place.
Corals may be solitary but most are colonial. Most are found in shallow waters; the accumulation of their calcium-carbonate remains builds reefs. Modern scleractinian coral, dominat reef-builders since the Triassic Period, have symbiotic photosynthetic dinoflagellates living withing the coral body. These dinoflagellates are in the genus Symbiodinium, and are termed zooxanthellae.
The polyp stage is dominant in members of this class. The Portuguese man-of-war is a colony of polyps, with the original polyp becoming a gas-filled float. Other polyps specialize for feeding or reproduction. The Portuguese man-of-war can cause serious injury to swimmers since each tentacle has numerous nematocysts.
Hydra are solitary, freshwater hydrozoan polyps. The body is a small tube about one-quarter inch in length, best observed with a dissecting microscope. Four to six tentacles surround the mouth, the only opening at one end. Hydra can move from one location by gliding or even somersaulting. Hydras have both muscular and nerve fibers, and respond to touch. Epidermal cells are termed epitheliomuscular cells and contain muscle fibers. Cnidocytes and sensory cells are also present in the epidermis. Interstitial cells can produce an ovary or testis, and may assist regeneration. Gland cells secrete digestive juices into the gastrovascular cavity into which tentacles have stuffed captured prey. Digestion is completed within food vacuoles of nutritive-muscular cells. Nutrients diffuse to the rest of the body. Hydras reproduce both sexually asexually (by budding). In sexual reproduction, sperm from a testis swim to an egg within an ovary. Following early development within the ovary, a protective shell forms and allows the embryo to survive until conditions are optimum. Hydra are commonly utilized animals in biology labs.
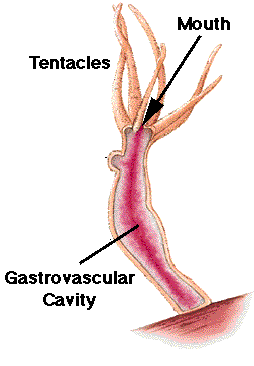
Anatomy of Hydra.
The medusal stage is dominant in jellyfish; the polyp remains small. Jellyfishes also serve as food for larger marine animals.

Image of the jellyfish Aurelia. Image from http://128.128.172.102/SPECIMENS/phylum.taf?function=detail&ID=29&branch=image. Photo illustrates the four circular gonads of the medusa phase, and the ring of tentacles at the rim of the medusa.
The fossil record of cnidarians is very good for corals, but usually not as good for soft-bodied forms like jellyfish. Corals become dominant reef-building animals during the Ordovician, and continue their importance today.
The Phylum Platyhelminthes contains about 13,000 species of flatworms subdivided into three classes: two parasitic and one free-living. The planaria and relatives are freshwater animals placed in the Class Turbellaria. Flukes are external or internal parasites belonginmg to the the Class Trematoda. Tapeworms are internal parasites and form the Class Cestoda. The phylum as a whole has adult bilateral symmetry and cephalization (the development of a head with sensory organs, in most members).
Flatworms have three tissue layers: ectoderm, mesoderm and endoderm and a body plan that is acoelomate and sac-like with a single opening. The mesoderm layer gives rise to muscles and reproductive organs. Free living forms have muscles, a nerve cord, and digestive organs, but lack both the respiratory and circulatory systems common to the "higher" animals. Flatworms have a branched gastrovascular cavity that is the site of extracellular digestion and which distributes nutrients throughout the body. Gas exchange occurs by diffusion through the skin. Platyhelminths have an excretory system that also functions as an osmotic-regulating system. Flatworms have ladder-style nervous system composed of paired ganglia that form a brain connected via nerve cells to sensory cells in the body wall.
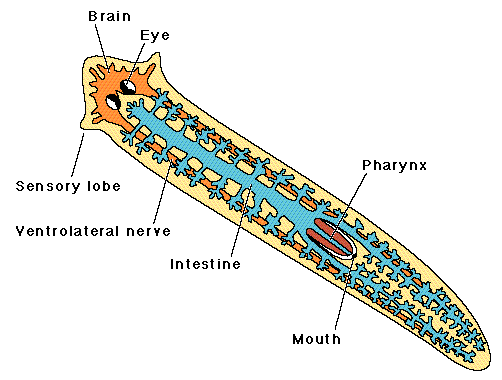
Anatomy of a flatworm. Note the sensory lobes and digestive system.
Parasitic members of this phylum, such as flukes and tapeworms, have characteristic modifications:
Both flukes and tapeworms use secondary or intermediate hosts to transport the species from primary host to primary host. The primary host is infected with the sexually mature adult while the secondary host contains the larval stage(s).
The Class Turbellaria includes freshwater planaria such as Dugesia that feed on small organisms or the remains of small creatures. Their small size and ease of care also make them common animals in biology labs. The planarian head is arrow-shaped, with side extensions that are sensory organs for detection of food and the presence of other organisms. Flatworms have two light-sensitive eyespots that have pigmentation making them look cross-eyed. The presence of three muscle layers facilitates varied movement. Gland cells secrete a mucous material upon which the animal slides or glides.
The animal captures food by wrapping itself around prey, entangling it in slime, and pinning it down. The pharynx is a muscular tube that extrudes from the mouth and through which food is ingested.
The flame cell system functions in excretion and consists of a series of interconnecting canals that run length of the body on either side of the longitudinal axis and side branches of the canals, each ending in a flame cell. The flame cell is a bulb-shaped cell containing a tuft of cilia within the hollow interior of the bulb. The cilia move back and forth, bringing water into the canals that empty through pores at the body surface. This flame-cell system functions in both water excretion and osmotic regulation in a typical freshwater, free-living flatworm.
Planaria can reproduce both sexually and asexually. They can constrict beneath the pharynx and each half will grow into a whole animal by the process of regeneration. Planaria are hermaphroditic, possessing both male and female sex organs, and can cross-fertilize each other. Fertilized eggs are enclosed in a cocoon and hatch in two to three weeks.
The Class Trematoda includes flukes. Flukes, such as blood, liver, and lung flukes are named after the organs they inhabit. Fluke bodies tend to be oval and elongate. They lack a definite head but have an oral sucker surrounded by sensory papillae. Flukes have reduced digestive, nervous, and excretory systems. Reproductive systems are well developed and usually hermaphroditic.
The blood fluke causes schistosomiasis, a disease found predominantly in tropical Africa and South America. Unlike most flukes, blood flukes are male or female. Flukes deposit eggs in blood vessels around the host's intestine. The eggs migrate to the intestine and are passed out with feces. Larvae hatch in water and swim about until they detect and enter a particular species of snail. The larvae reproduce asexually and eventually leave the snail. Once larvae penetrate human skin they begin to mature in the liver, and implant in blood vessels of the small intestine. A weakened person is then more likely to die from secondary diseases.
The Chinese liver fluke requires two intermediate hosts. Humans become infected when they eat uncooked fish. Adults migrate to the liver and deposit eggs in the bile duct, which carries the eggs to the intestine. The larval flukes must then pass through two intermediate hosts, a snail and a fish.
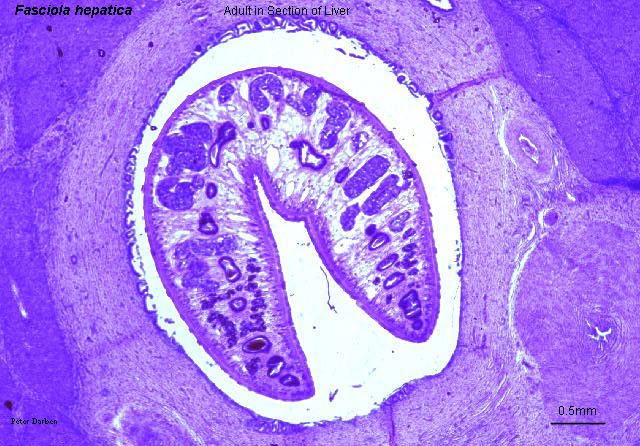
Fasciola hepatica, a liver fluke, in a section of liver, from http://www.life.sci.qut.edu.au/LIFESCI/darben/figs/flukes/fhepasec.jpg, a page at The Parasitology Images List by Peter Darben.
The Class Cestoda consists of the tapeworms. The tapeworm scolex has hooks and suckers that allow the organism to attach to the host's intestinal wall. Behind the head is a short neck and then a long string of proglottids. Each proglottid segment contains a full set of both male and female sex organs and very little other structure. Since the animal does produce wastes, it retains its excretory canals, but no digestive system is needed. Being sessile organisms in a fairly stable environment, tapeworms have only rudiments of nerves. Following fertilization, proglottids become a bag of eggs that when mature, breaks off and passes out with feces. If the eggs of tapeworms are ingested by pigs or cattle, the larvae become encysted in the muscle of the hosts. The covering of ingested eggs is digested away and the larvae burrow through the intestinal wall and travel by bloodstream to lodge and encyst in muscle; a cyst is a hard-walled structure sheltering a larval worm. If humans eat the meat of infected pigs or cattle and fail to cook it properly, they too become infected.
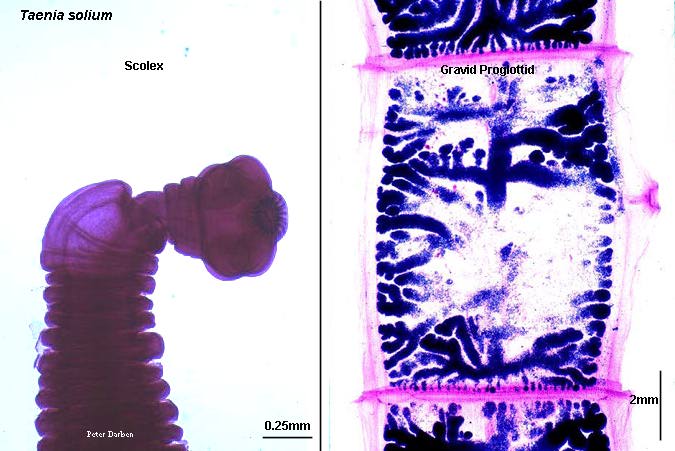
Taenia solium scolex and gravid proglottis, from http://www.life.sci.qut.edu.au/LIFESCI/darben/figs/tapeworm/tsoliscp.jpg, a page at The Parasitology Images List by Peter Darben.
The Phylum Nemertea include approximately 650 species of marine ribbon worms. Ribbon worms have a distinctive eversible proboscis stored in a rhynchocoel; when the walls of the rhynchocoel are contracted, the proboscis is extended out of the body. The proboscis is a long, hollow tube that can be everted and shot outward through a pore located just above the mouth. It is used primarily for prey capture, and for defense, locomotion, and burrowing. This phylum is included as the oranisms are also triploblastic. Several of the fossils from the Cambrian-aged Burgess Shale are interpreted as ribbon worms, and some extremely long worms have been found in the Mediterranean (up to 100 feet long) and in the ocean under the Antarctic ice shelves.
Rotifers belong to the phylum Rotifera, which contains about 2,000 species. Rotifers are often observed in biology lab preparations. Their digestive tract is the inner tube and the rest of the animal is the outer tube. Rotifers are microscopic and abundant in freshwater. A crown of cilia (corona) forms a rotating wheel that serves as both an organ of locomotion and acts to deliver food to the mouth.
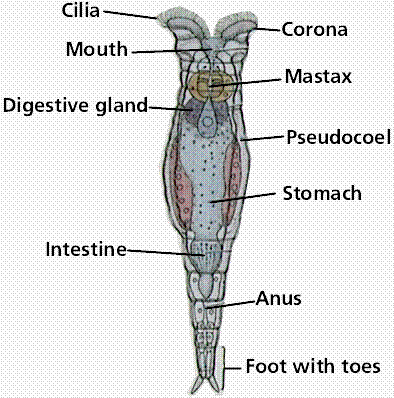
Anatomy of a typical rotifer. These microscopic animals are commonly seen in lab aquaria and cultures, but still exhibit the tube-with-a-tube body plan.
The Phylum Nematoda consists of several hundred thousand species of roundworms. Most are free-living, although some are parasitic (pinworms are thought to infect 30% of all US children). Adult nematodes have a pseudocoelom (tube-within-a-tube), a closed fluid-filled space that acts as a hydrostatic skeleton, aids in circulation and dispersal of nutrients. Nematodes lack a circulatory system, but do have a well developed digestive system.
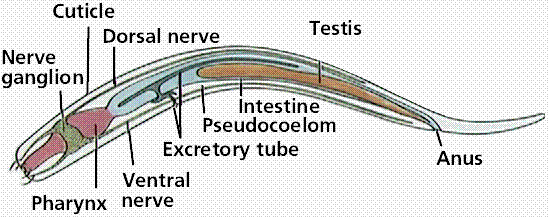
Anatomy of a nematode. Nematodes, like rotifers, are pseudocolelomates.
Image of Tylocephalus auriculatus, a nematode, from http://allserv.rug.ac.be/~pdeley/tyloceph.html, a part of A Gallery of Weird Worms.
One nematode, Caenorhabditis elegans, has only 1000 genes in its genome and its developmental pathways are well known. C. elegans serves as a model for eukaryote gene systems and has been extensively studied as part of the human genome project.
Ascaris is a parasitic roundworm. These worms are unsegmented and have a smooth outside wall. They move by whiplike motions. Mating produces eggs that mature in the soil, limiting most are to warmer climates. When eggs are swallowed, larvae burrow through the intestinal wall, moving to the liver, heart and/or lungs. Once within the lungs, larvae molt and, after 10 days, migrate up the windpipe to throat where they are swallowed. In the intestine, the mature worms mate and the female deposits eggs that are lost with the feces. Feces must reach the mouth of the next host to complete the life cycle, thus, proper sanitation is an important aspect to prevent infection.

Ascaris lumbricoides, female (top) and male (bottom), from http://www.life.sci.qut.edu.au/LIFESCI/darben/figs/nematode/intnema/alumbmf.jpg, a page at The Parasitology Images List by Peter Darben.
Humans contract Trichinella (which causes the disease trichinosis) by eating raw pork containing encysted larvae. Mature female adults burrow into the wall of the small intestine. Live offspring are carried by the bloodstream to the skeletal muscles where they encyst. Religious dietary injunctions against eating pork may in part be a reflection of the prevalence of this disease in the Middle East.

Trichinella spirallis, from http://www.life.sci.qut.edu.au/LIFESCI/darben/figs/nematode/othnema/tspirlrv.jpg, a page at The Parasitology Images List by Peter Darben.
Filarial worms cause various diseases. Dirofilaria causes heartworm in dogs, and is a common filarial worm in temperate zones.
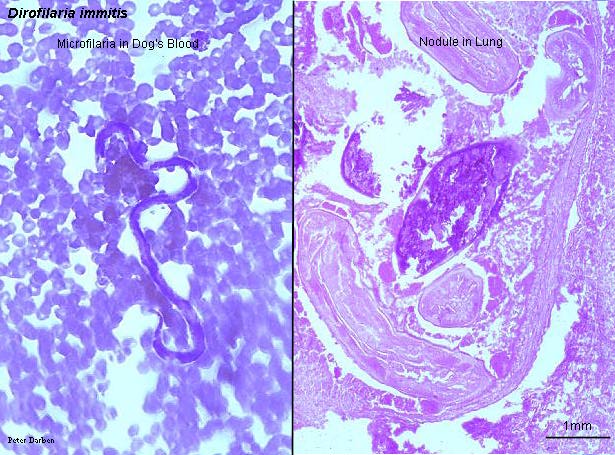
Dirofilaria immitis, a filarial worm, in dog's blood and lung, from http://www.life.sci.qut.edu.au/LIFESCI/darben/figs/nematode/othnema/dimitfsc.jpg, a page at The Parasitology Images List by Peter Darben.
Elephantiasis, a disease associated with tropical Africa, and is also caused by a filarial worm that utilizes mosquitos as secondary hosts. Adult worms reside in and block lymphatic vessels. This results in limbs of an infected individual swelling to monstrous size. Elephantiasis is treatable in its early stages but not after scar tissue has blocked lymphatic vessels.
Reitner, J. 1990. Polyphyletic origin of the "Sphinctozoans". In Rutzler, K. (ed.), New Perspectives in Sponge Biology, Proceedings of the Third International Conference on the Biology of Sponges (Woods Hole). pp. 33-42. Smithsonian Institution Press, Washington, DC.
Email: mj.farabee@emcmail.maricopa.edu![]()
Last modified:
2000/01/05:09:53:37
The URL of this page is:
gened.emc.maricopa.edu/bio/BIO181/BIOBK/BioBookDiversity_7.html
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}