![]()
![]()
![]()
![]()
![]()
![]()
![]()
| Topics covered on this page are:
|
 |
Long before Charles Darwin published "The Origin of Species" people had tried to explain the diversity of living things they observed around them. Theologians had a nice, convenient story of creation, and for many thousands of years that belief satisfied almost everyone. (If it's still good for you, that's fine, but please keep on reading, beacuse you will not get marks for exam questions with a creationist answer.)
But, as always, there were the sceptics who needed scientific 'proof' and set about forming and testing their own hypotheses. One of the popular explanations for species diversity around Darwin's time came from Jean-Baptiste Lamarck. Lamark's theory of acquired characteristics was so popular that it is today often known as 'Lamarkism', but scientists can find no evidence to support this thoery any more. (You should read a little about Lamarkism from a biology text, just to know how NOT to answer exam questions.)
Whilst reading of "The Origin of Species" is probably not high on your priority list at present, I'd recommend that if you're seriously interested in understanding where the planet's species, including our own, came from, it is a fascinating piece of writing and scientific deduction. In brief, Darwin, during his journey around the world on The Beagle made detailed observations of the animals and to a lesser extent the plants he found. On his return to England, he pondered these observations for many years, trying to make sense of them and explaining the patterns he had observed. It was many years before he published the book which still underpins what scientists now believe about species, their evolution and their relationships.
Here is an outline of Darwin's observations (O) and deductions (D):
(As you read this list think about a few species other than humans - we tend not to obey Darwin's "rules" - with consequences which may prove to be fatal!)
Add to Darwin's theory another 100+ years of scientific investigation, especially the advances in molecular biology techniques of the recent past, and we have a story that's difficult to dismiss:
The origins of variation within a population are mutation and random assortment of chromosomes during meiosis. These mechanisms, then, can be considered as the genetic basis for natural selection.
Without variation within a population, there could be no natural selection - the population would either survive or die as minor environmental changes were encountered, since all members of the population would be genetically identical and all would be equally well or badly equipped to deal with the changes.
Natural selection can be described as the varying success in reproduction of different phenotypes resulting from their interaction with the environment. Therefore, unless there are different phenotypes in a population, there can be no natural selection.
There are three common ways that natural selection can act:
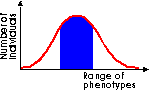 |
Stabilising selection removes the extreme variants from a population, reducing the trend towards phenotypic variation and maintaining the status quo. This is most likely to be the case where an environment is itself stable. |
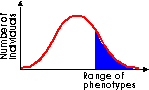 |
Directional selection shifts the overall makeup of the population by favouring one extreme phenotype. This is likely to occur when the environment undergoes a marked change. The case study of the Biston betularia (peppered moth) illustrates this. (Look this up in your textbook, it's almost certain to be there.) |
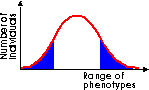 |
Diversifying selection favours the extremes over intermediates. If the environment of the species contains two different features (eg. in background colour - light/shade) two different phenotypes have a selective advantage, depending on location. |
 |
Another form of natural selection occurs in species where the males and females are very different in appearances or behaviour, apart from the physiological differences due to gender. This is called sexual dimorphism, an example of which is plumage (feather colour) differences in male/female birds. This can itself, be an agent of natural selection, with the best plumed male birds gaining the largest number of matings within their population. This ensures that the genes for the "desirable" traits are passed on to the next generation and that genes from the "less desirable" males are reduced or eliminated. |
The gene pool can be defined as the total aggregate of genes, and all of their alleles, in a population at any one time. For a diploid individual, each gene locus is represented twice in the genome, and the two alleles may be the same (homozygous) or different (heterozygous). There may, however, be more than two alleles for a gene within a population's gene pool (eg the ABO blood groups in humans), although each individual can only have two of those alleles.
If a population of a species is stable and non-evolving, the composition of that population's gene pool is constant, since it is in equilibrium. This is known as the Hardy-Weinberg Theorem and it can be demonstrated mathematically to be true. It is stated as follows:
| No matter how many generations' alleles are segregated by meiosis and combined by fertilization, the frequencies of the alleles in the gene pool will remain constant unless acted on by other agents. |
For this to be true, five conditions must be met:
Whilst these conditions can be approached in nature, they are never achieved for very long.
Population genetics is the scientific study of gene pools and the genetic variation within populations.
Because the five conditions for Hardy-Weinberg equilibrium are seldom maintained for many generations, gene frequencies in gene pools do change.
If a population is small, genetic drift may occur. Chance events in matings may cause the allele frequencies to drift randomly from generation to generation. (eg Toss a coin 10 times, you will not necessarily get 55 heads and 5 tails.) Only luck would result in these random changes improving adaptations.
Sometimes non-representative sub groups of a larger population become isolated. This can lead to the founder effect, where a new, different allele frequency exists in the founding population and is maintained in subsequent generations. This can be seen in some small isolated populations of humans where the frequency of inherited diseases is high and can be traced back to an original founder of the settlement.

|
The bottle-neck effect is similar to the founder effect. It can be seen when a population is reduced to a very small number, such as by natural disaster or recently by unnatural activity imposed by humans (eg hunting of gorilla to near extinction). The allele frequencies in the surviving population may not reflect the original larger population so phenotypic variation may be reduced. |
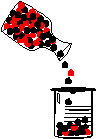
|
This is illustrated in the diagram at left. The red and black balls represent the two alleles of a particular gene. They are present in approximately equal numbers in the original population. In the isolated population (the beaker), chance has caused more black balls than red balls to be present. If, sometime in the future, red balls (alleles) would have given a survival advantage, there may not be enough of them in the population to prevent extinction. |
A phenotype with an environmentally poor survival outcome is less likely to be maintained in a population, and the frequencies of the alleles which code for this phenotype would be expected to decrease in the population's gene pool.
Since most populations are not entirely reproductively isolated, the population may gain or lose alleles by gene flow, the migration of fertile individuals or the transfer of gametes, between populations.
A new mutation that is transmitted to gametes must immediately change the gene pool by substituting one allele for another. This is true whether or not the mutated allele is "new" to the gene pool, although the effects of a mutation are likely to be more pronounced if it gives rise to a new phenotype - better or worse.
Nonrandom matings are more common that one might first expect. In many populations, family groups stay within loose geographical boundaries within the larger group. This can lead to localised inbreeding and pockets of very different allele frequencies within the larger group. Similarly, if individuals with a particular phenotype are perceived as more desirable mates (eg Elle MacPherson compared with Elle MacFeast or James Hird compared with John Howard (sorry if you're not Australian, this is 'local humour'!)), the "desirables" (assuming that they are willing to mate with multiple partners!) are more likely to pass on more of their alleles to the next generation. This is termed assortative mating.
Similarly, a genotype which leads to a phenotype with a positive survival advantage in the face of a changing environment would be expected to increase in a population's gene pool over many generations. This is the basis of natural selection.
All species have such great potential fertility that their population size would increase exponentially if all individuals born would eventually reproduce, at their full potential. The fact is that populations are generally relatively stable in size, except for seasonal variation.
What keeps populations constant? Why do some survive? Why do many die? The answers to these questions all help explain the struggle for survival which occurs constantly in all populations of all species, including our own.
| Selection pressures are those biotic and abiotic factors of the environment that can influence the survival and reproduction of individuals, and ultimately of populations and species. |
Selection pressures include:
Individuals with structural (anatomical), functional (biochemical) or behavioural adaptations which give them a better chance of survival to adulthood and/or give them a reproductive advantage are more likely to pass their genes on to the next generation of the population.
NOTE: The correct term is 'adaptation' , not 'adaption' please try to remember this.
Adaptations are acquired by an individual as a set of genetic information - the chance combination of alleles obtained at fertilization.
Avoid the notion that "individuals can adapt" - This is Lamarckism and is not able to be supported scientifically. (Read about Lamark to understand this fully)
Individuals can only express in their phenotype the characteristics for which their genotype codes. This can include many variants of phenotype, within the one individual. For example a weight lifter changes his body shape over time with a great deal of training, but his children will not be born as mini-Schwarzeneggers because he did so! What the weight lifter has done is manipulate his own environment to realise all his genetic potential in his phenotype.
The phrases "struggle for survival" and "survival of the fittest" can be misleading if taken literally.
They are not meant to mean that only the strongest, physiologically fittest individuals will survive and reproduce. Rather they mean that species will acquire favourable characteristics (become "fitter") which enable viable populations of that species to survive over many generation.
Relative fitness is the contribution of a genotype to the next generation compared to the contribution of alternative genotypes at the same locus. From this it can be seen that the phenotype corresponding to a high frequency genotype is probably advantageous in the environment of the species at the time - or at least it is not disadvantageous.
| Natural selection is the differential success in reproduction of different phenotypes resulting from interaction of organisms with their environment. With time, natural selection causes changes in relative frequencies of alleles in the gene pool. |
| Species = A particular kind of organism; members of a species possess similar anatomical characteristics and have the ability to interbreed, producing vigorous (= healthy), virile (= able to reproduce sexually) offspring. |
When a population of a species splits, usually due to selective pressures as previously described, the populations may become reproductively isolated. As described earlier, this will almost certainly, over many generations, lead to changes in the gene pool of each sub-group. As these changes progress, random chance makes it highly likely that significant differences in phenotypes will develop in each group.
For a while, breeding between groups will remain theoretically possible, if individuals were to meet and mate. Eventually, however, the accumulated changes in each group's gene pool would lead to a significant lack of homology at a number of gene loci. Viable offspring between the groups would now be impossible - the groups are no longer members of the same species - speciation has occurred.
(Whether there is one new species and the original species in the other group, or whether there are two new species, both different to the original, could only be determined by observations of outcomes of mating experiments. Can you work out how a biologist might investigate this?)
Speciation has been, and still is, occurring constantly since life began on Earth. We can see evidence of it in nature. For example, Australian magpies, species Gymnorhina tibicen, show phenotypic variation in different populations on the east coast, from Tasmania to Cape York. In some of these sub-groups (sometimes termed races), interbreeding is still possible, especially if their home ranges overlap. Where the populations are geographically separated by large distances or high mountains, the populations may no longer able to successfully interbreed. Deciding exactly where one species stops and another starts can be very difficult in cases such as this.
Human activity has recently hastened this process by manipulating selection criteria in many species. Many cultivated plants, bred originally from natural "wild" species bear little resemblance to the originals and are now infertile, being reproduced asexually (eg by cuttings) for commercial purposes.
Sometimes environments change relatively rapidly. If the species which live in those changing environments are not lucky enough to already possess genes which enable the species to survive in the face of such change, or if random mutations do not accumulate sufficiently quickly, all members of the unlucky species may die. This is extinction.
Extinction may be localised to one species in an ecosystem, often due to destruction of that species habitat, or it may widespread.
The fossil record indicates that there have been at least twelve periods during the history of life on Earth where mass extinctions occurred. The two most extreme of these occurred about 250 million years ago (>90% of marine species died) and 65 million years ago (>50% of marine species, and many terrestrial species, including the dinosaurs died). Paleobiologists are still divided as to the explanations for these, asteroid collisions and global climate changes are currently the two leaders in the explanation stakes.