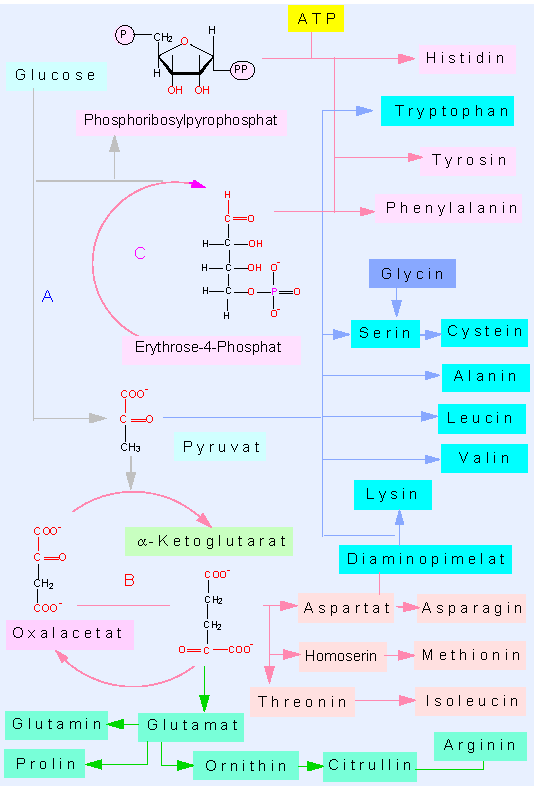
Pflanzen sind in der Lage, alle 20 für den Aufbau von Proteinen benötigten Aminosäuren selbst zu synthetisieren. Darüber hinaus bilden sie eine Reihe weiterer, von denen wir einige im folgenden Abschnitt behandeln werden.
Bei der Besprechung der Glykolyse und des Citratzyklus hatten wir es mit kohlenstoffhaltigen, doch stickstofffreien Molekülen zu tun gehabt. Von einigen der Intermediärprodukte (Zwischenprodukte) zweigen Syntheseketten der Aminosäuren ab. Aufgrund chemischer Gemeinsamkeiten - und nur weniger Ausgangssubstanzen - können wir sie fünf Gruppen (Familien) zuordnen:
- Glutamatfamilie (ausgehend vom alpha-Ketoglutarat)
- Aspartatfamilie (ausgehend vom Oxalacetat)
- Alanin-Valin-Leucin-Gruppe (ausgehend vom Pyruvat)
- Serin-Glycin-Gruppe (ausgehend vom 3-Phosphoglycerat)
- Aromatische Aminosäuren (ausgehend vom Phosphoenolpyruvat und dem Erythrose-4-Phosphat).
Woher stammt nun der Stickstoff? Pflanzen nehmen ihn in Form von Nitrat, in geringem Umfang auch in Form des Ammoniumions auf. Einige wenige Arten, vorwiegend Leguminosen, leben in Symbiose mit stickstoffbindenden Bakterien, die zur Reduktion atmosphärischen Stickstoffs befähigt sind. Die übrigen Pflanzen begnügen sich mit einer Nitratreduktion.
Nitrat wird in einem ersten Schritt zu Nitrit reduziert. Die dafür benötigte reduzierende Substanz ist das NADH + H+, das bei der Glykolyse anfällt, das erforderliche Enzym ist die Nitratreduktase. Damit wären Glykolyse und Nitratreduktion miteinander gekoppelt, was unter anderem auch mit dem Vorteil verbunden ist, daß regeneriertes NAD+ für den Fortgang der Glykolyse anfällt.
In einem zweiten Schritt muß Nitrat zum Ammoniumion weiterreduziert werden. Als Elektronendonor wirkt dabei reduziertes Ferredoxin, das seinerseits im Photosyntheseprozeß gewonnen wird. Der hierfür benötigte Enzymkomplex ist die Nitritreduktase, die in photosynthetisierenden Geweben in den Chloroplasten lokalisiert ist.
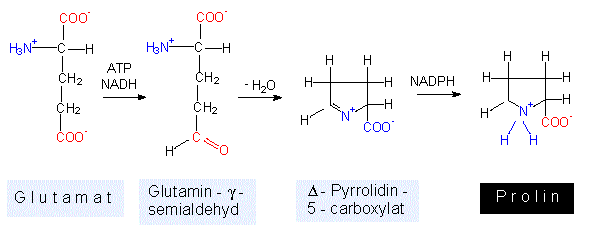
Die Reaktion läuft über eine Elektronentransportkette, an der sowohl NADP als auch FAD beteiligt sind. Wie die Schritte im einzelnen zusammenhängen, ist zum Teil noch ungeklärt. Freie Ammoniumionen sind für jede Zelle toxisch, sie müssen daher schnellstmöglich abgefangen werden. Der quantitativ wichtigste Weg dürfte bei grünen Pflanzen über die NADP+ - abhängige Glutamat-Dehydrogenase-Reaktion laufen. Damit wären wir bereits bei der ersten Aminosäure und hätten einen Einstieg zur Besprechung der Glutamatfamilie. Zuvor sollte noch eine zweite Alternative genannt werden: Das Glutamat selbst kann ein weiteres Ammoniumion binden und wird so in die Aminosäure Glutamin überführt. Auch diese Reaktion läuft in den Chloroplasten ab. Beide Reaktionen sind ATP-abhängig.
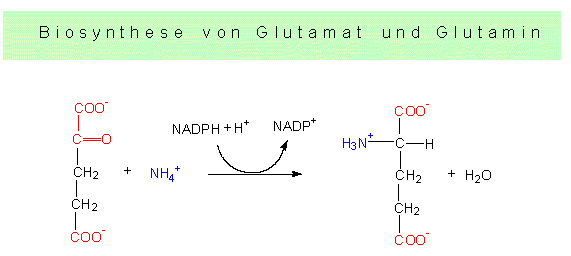
Glutamatdehydrogenasereaktion: Ausgangsprodukte sind alpha-Ketoglutarat und NH4+
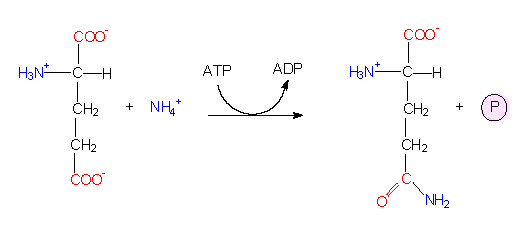
Glutaminsynthetasereaktion: Ausgangsprodukte sind Glutamat und NH4+
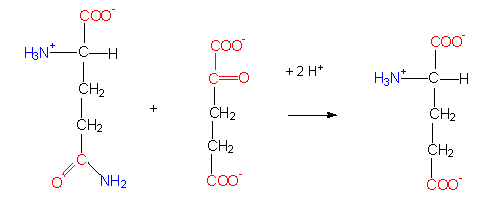
Glutamatsynthasereaktion (eine Transaminierung). Die Aminogruppe einer Aminosäure wird auf eine alpha-Ketosäure (hier: alpha-Ketoglutarat) übertragen.
Zum Verständnis von Aminosäuresynthesen ist der Einfluß
einer Gruppe von Enzymen, der Transaminasen, hervorzuheben. Diese sind
in der Lage, eine Aminogruppe (vorwiegend) von Glutamin auf eine alpha-Ketosäure
zu übertragen. Damit kommt ihnen gleichsam eine Verteilerrolle von
Aminogruppen zu. Keimende Samen, denen funktionsfähige Chloroplasten
fehlen und die daher keine Photosynthese betreiben, beziehen Aminogruppen
ausschließlich aus dem Abbau vorhandener Reserveproteine und schleusen
sie auf dem eben skizzierten Weg in neu zu bildende Aminosäuren, Proteine
sowie weitere stickstoflhaltige Verbindungen ein.
Die Glutamatfamilie. Hierzu gehören die Aminosäuren Glutaminsäure (Glutamat), Glutamin, Prolin und Arginin. Die Synthese der ersten beiden kann auf unterschiedlichen Wegen erfolgen
Prolin entsteht durch Ringschluß - über zwei Zwischenstufen - unter Verbrauch von je einem ATP, NADH + H+ und NADPH + H+. Das weist darauf hin, wie energieaufwendig Biosynthesen sind. Im folgenden wird das nicht jedesmal explizit erwähnt, doch sollte man sich vergegenwärtigen, daß jede vergleichbare Biosynthesekette Energiebeträge dieser Größenordnung verbraucht. Prolin enthält keine Aminogruppe, korrekterweise dürfte man es daher nicht als Aminosäure, sondern müßte es als Iminosäure bezeichnen (deren Merkmal: >NH).
Eine Umwandlung von Prolin in Hydroxyprolin erfolgt nur an Prolinresten, die Bestandteil einer Polypeptidkette sind. Auffallend häufig ist Hydroxyprolin in einem Zellwandprotein, dem Extensin.
Arginin ist eine basische Aminosäure, d.h., sie trägt eine zweite, unter physiologischen Bedingungen ionisierte Aminogruppe. Ihre Biosynthese läuft über insgesamt acht Zwischenstufen. Ein wichtiges Zwischenprodukt ist Ornithin, das in pflanzlichen Zellen auf zwei alternativen Wegen gebildet werden kann. Es dient als Akzeptor für Carbamylphosphat. Durch dessen Anlagerung, bei gleichzeitiger Phosphatabspaltung, wird die Molekülkette des Ornithins verlängert, und es entsteht Citrullin.
Beide Zwischenprodukte gehören in diejenige Kategorie von Aminosäuren, die zwar in Zellen häufig vorkommen, doch nicht in Proteine eingebaut werden. Unter Anlagerung von Aspartat wird das Molekül erneut vergrößert. Aus dem gebildeten Produkt wird dann ein Teil wieder herausgespalten (Fumarat), wobei Arginin entsteht. Das Beispiel dieser Biosynthese macht mehreres deutlich:
| Die Biosynthese komplexer Produkte läuft über eine Vielzahl von Zwischenprodukten. | |
| Durch Zusammenlagerung zweier Moleküle wird die -C-C- Kette verlängert. Es wird also nicht jede Bindung für jedes Produkt neu geknüpft. | |
| Aminosäuren werden außer zum Einbau in Proteine auch als Zwischenprodukte für die Synthese anderer Aminosäuren benötigt. | |
| Biosynthesewege sind untereinander vernetzt. |
Die Aspartatfamilie. Zu dieser Gruppe rechnet man die Aminosäuren Asparaginsäure (Aspartat), Asparagin, Threonin, Isoleucin und Methionin. Aspartat entsteht aus der Transaminasereaktion
Glutamat + Oxalacetat > alpha-Ketoglutarat + Aspartat
und Asparagin durch Aufnahme einer weiteren Aminogruppe. Das kann einmal durch die Fixierung eines Ammoniumions geschehen (analog der Bildung von Glutamin), zum anderen aber auch durch eine Übernahme einer Aminogruppe vom Glutamin.
NH4+ + Aspartat + ATP > Asparagin + ADP + Pi
Glutamin + Aspartat + ATP > Glutamat + Asparagin + AMP + PP
Vom Aspartat führt ein Weg zum L- Homoserin, einem Zwischenprodukt (Intermediärprodukt), von dem aus sich die Wege zum Threonin und Isoleucin einerseits und zum Methionin andererseits trennen. Die Reaktionskette zum Threonin führt wieder über mehrere Zwischenprodukte, wobei zwei ATP-Moleküle, ein NADPH + H+ und ein NADH + H+ verbraucht werden. Es gäbe eine energetisch günstigere Lösung:
Acetaldehyd + Glycin > Threonin
doch ist Acetaldehyd in aerob lebenden Zellen Mangelware (abgesehen davon, daß es ein starkes Zellgift ist).
Die Synthese des Isoleucins geht vom Threonin aus, für die des Methionins wird Schwefel benötigt. Dieser wird von Pflanzen als Sulfation aufgenommen und in einer Zweischrittreaktion in die reduzierte Form (formal Schwefelwasserstoff) gebracht. Sulfid entsteht als Zwischenprodukt. Ein für die Reaktion benötigtes Enzym konnte aus Chloroplasten (von Spinat) isoliert werden. Wiederum wird das reduzierte Ferredoxin als Elekronendonor benötigt. Der reduzierte Schwefel erscheint letztlich als -SH-Gruppe im Cystein (assimilatorische Sulfatreduktion).
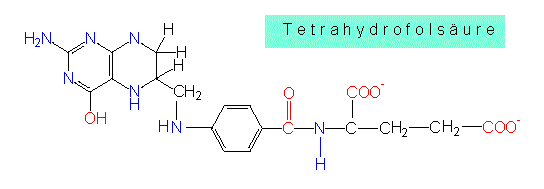
Von dort wird sie auf eine Vorstufe des Methionins übertragen. Andererseits gibt es aber auch einen Weg, bei dem Cystein aus Methionin wiedergewonnen werden kann. An der Methioninbildung ist ein Chlorid-Gruppen-Überträger, die Tetrahydrofolsäure (THF), beteiligt. In diesem Fall überträgt sie eine Methylgruppe und koppelt sie an freies -SH. Ergebnis: -S-Methyl.
Lysinsynthese. Wie bei der Kopplung zwischen Arginin- und Asparaginsynthese, findet man auch eine Kopplung zwischen Produkten der Glutamat- und der Aspartatfamilie. Lysin kann auf zweierlei Weise entstehen. Die beiden Wege sind nach den charakteristischen Zwischenstufen benannt. Bei Grünalgen, Farnen und höheren Pflanzen wird der erste Weg beschritten, während bei einigen grünen Flagellaten (Euglenophyta) sowie bei Pilzen der zweite Weg eingeschlagen wird.
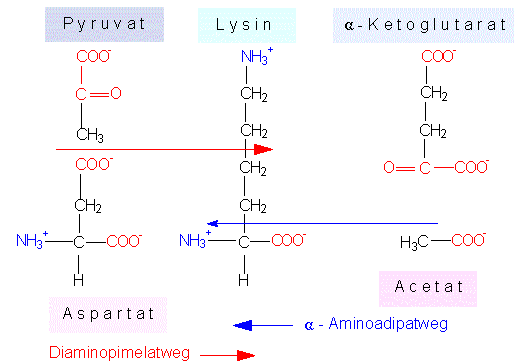
Lysinsynthese auf zwei Wegen. Beim Diaminopimilatweg entsteht das Kohlenstoffgerüst des Lysins aus Pyruvat und Aspartat, beim alpha-Aminoadipatweg aus alpha-Ketoglutarat und Acetat.
Alanin-Valin-Leucin-Gruppe: Hier sind die Verhältnisse wieder übersichtlicher. Ausgangsprodukt ist das Pyruvat, dessen Aminoderivat das Alanin ist. Valin und Leucin entstehen unabhängig voneinander durch Kettenverlängerung des Pyruvats, Umlagerung und anschließende Transaminierung.
EINE ZUSATZBEMERKUNG: Leucin und Isoleucin sind von ihrer chemischen Struktur und ihren Eigenschaften her nahe verwandt, ihre Synthesewege sind jedoch grundsätzlich voneinander verschieden.
Serin-Glycin-Cystein-Gruppe.
Serin wird in einer Zweischrittreaktion aus 3-Phosphoglycerat gebildet.
Der erste Schritt ist eine Oxydation, der zweite eine Transaminierung.
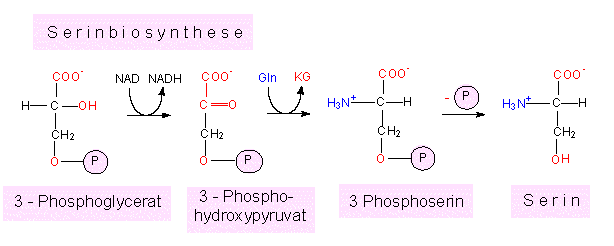
Alternativ dazu kann 3-Phosphoglycerat dephosphoryliert werden, wobei Hydroxypyruvat entsteht, das seinerseits nach Transaminierung ebenfalls zu Serin führt.
Vom Serin führt ein Weg zum Glycin, wobei die Seitenkette um den Anteil -CH2OH verkürzt wird und die C1-Einheit von der bereits genannten Tetrahydrofolsäure übernommen wird. Ein weiterer Weg führt zum Cystein. Das macht deutlich, daß auch Cystein auf mindestens zwei Wegen (der andere war Abbau von Methionin) entstehen kann.
Aromatische Aminosäuren: Außer dem Phosphoenolpyruvat wird Erythrose-4-Phosphat als Ausgangsprodukt benötigt. Über einige Zwischenstufen kommt es zu einer Ringbildung, wobei zunächst das Shikimat, und nachfolgend das Chorismat, als wichtige Zwischenprodukte zu nennen sind. Letzteres ist Ausgangsprodukt für drei getrennte Wege, die zu den Endprodukten Phenylalanin, Tyrosin und Tryptophan führen können.
Für die Synthese jeder dieser Aminosäuren sind verschiedene Wege möglich. Bei der Tryptophansynthese greift eine aktive Form des Ribosephosphats (Phosphoribosylpyrophosphat) ein. Es spielt auch bei der Synthese des Histidins (die hier nicht besprochen wird) sowie der Purine und Pyrimidine eine Schlüsselrolle.
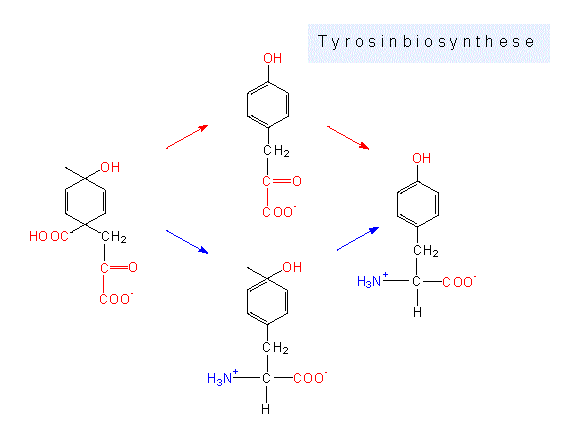
Alternative Biosynthesewege. Die hier als Ausgangssubstanz gezeigte Verbindung ist ein labiles Zwischenprodukt, das unter Mitwirkung der Chorismat-Mutase aus Chorismat entsteht.
|
|
 Prolin
Prolin